


2017 27.2 Biodiversity and Natural Pest Suppression in the Pacific Northwest
Maintaining biodiversity on the farm is something many of us strive for in order to promote stability and resilience in the agroecosystem. At WSU, we have tackled research questions related to how biodiversity supports natural pest suppression for almost 15 years, and we have come a long way in understanding how to harness the benefits of diverse beneficial insect communities.
We specialize in the diversity of predatory insects like lady beetles, assassin bugs, parasitoid wasps, ground beetles and lacewings, which prey on herbivores like aphids, caterpillars and flea beetles. The idea that diversity begets pest suppression started in the 1970s, with the “Enemies Hypothesis.” This hypothesis suggests that diverse plant communities support predator diversity by providing diverse supplemental food and habitat resources1. If you’re interested in resources to help decide what kinds of plants to include in your crop areas, check out the “conservation biological control” page on the Xerces Society website; it’s the best clearinghouse of information out there.
 |
||
| Researcher Jake Asplund uses the “Dvac” to sample insects in a field of broccoli. |
Did you know that most predators of pests in agroecosystems are actually omnivores? It turns out that pests are not a particularly reliable food source, so it’s feast or famine for predators without diverse diets. Lots of evidence supports the theory that plant diversity supports predator diversity, and there are a number of easy ways to squeeze in a little more plant diversity on the farm by incorporating cover crops or insectary plants that provide nectar and pollen throughout the year.
So, increasing plant diversity is a great way to increase predator diversity, but the link between predator diversity and pest suppression is not quite so simple. Theoretically, diverse predator communities should provide stronger pest suppression because they are more likely to fill all the “trophic niches,” or parts of the food web, in the system. That means that the more types of predators there are, the more likely you are to find ones that eat each of the various pests you struggle with. For example, lady beetles mostly eat aphids, and other predators might feed more frequently on caterpillars. When all the predators have feeding preferences that complement each other, that tends to work great for biological control. However, sometimes predators eat other predators! For this reason, predator diversity can occasionally have a negative effect on pest suppression. For example, parasitoid wasps, which lay their eggs inside other insects, are extremely effective at controlling aphids, but they are often eaten inadvertently by lady beetles before the baby parasitoid has the chance to mature and emerge from the dead aphid carcass. If lady beetles kill enough parasitoids, they may eventually cause a reduction in biological control compared to what the parasitoid might have achieved alone.
So, we have examples of predator biodiversity being both good and bad for pest suppression, and it’s a major challenge to identify the type of predator community that provides the best ecosystem services. We have found that it’s not just predator diversity that drives pest control per se, but predator community “evenness” that leads to the strongest suppression2. This just means that the predator community is balanced, and not dominated by a single aggressive, voracious predator that might be more likely to attack the other predators in the agroecosystem. It turns out that predator community evenness is a unique trait that distinguishes organic farms from conventional ones. So, something about organic farms makes them more likely to support communities of predators that spend more time eating pests than eating each other. Stronger rates of natural pest control are frequently reported when conventional and organic farms are compared side-by-side, and this “community evenness” concept could explain that pattern. We’re still not sure exactly what it is about organic farms that supports balanced predator communities, but we have some promising ideas that we’ve been testing, related to non-crop plant resources and non-pest prey, described below.
Non-Crop Plant Resources
This idea hearkens back to the plant diversity concept from the “enemies hypothesis.” If multiple predators are hunting on the soil surface, they may be less likely to encounter each other if they’re hunting under a canopy of living mulch, or even some light weed growth. The structural complexity provided by the supplemental plant material might serve as a visual and structural buffer for small, active-hunting predators at risk of being eaten by larger ones, but should not hinder them as they find and kill sedentary pests3. In addition to providing refuge, many non-crop plants provision food resources, like nectar and pollen.
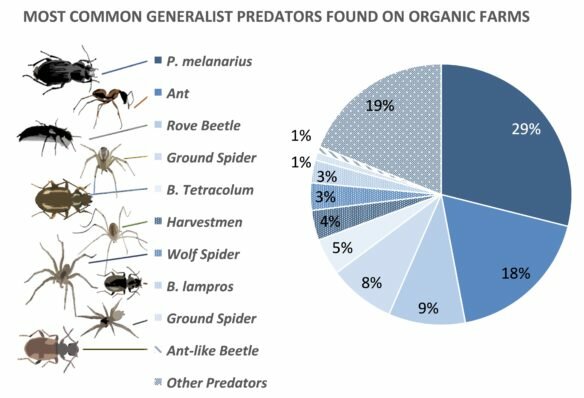 |
||
| Figure 1. Composition of the predatory insect communities sampled at the soil surface on more than 50 farms in Washington, Oregon and Idaho. |
Non-Pest Prey
Predator community evenness might also be conserved in systems that support a healthy stock of “alternative prey.” Springtails, for example, are decomposer arthropods, and pose no risk to crops, but may serve as an excellent food resource for predators during times of pest scarcity. Alternative prey allow predators to persist in crop areas in the absence of pests, and may also keep predators from feasting on each other. So, by serving as a stable food source, arthropods like springtails could indirectly lead to stronger suppression of pests once their populations start to build.
For the past two years, a group of ecologists in the Entomology Department at WSU have been on a mission to identify the particular qualities of organic farms that support predator diversity and predator community evenness. We’ve started by performing a huge survey of predator communities across more than 50 organic farms in Idaho, Washington, and Oregon. We have adopted a nomadic lifestyle each summer, and spend our days counting aphids and caterpillars on brassica crops.
To survey the predator community at the soil surface, we use pitfall traps–a pitfall trap is just a plastic cup sunk into the ground that captures unsuspecting predators who fall into the pit (see Figure 1 for the most common predators found in our survey). We sample predators in the foliage using a “Dvac” (the D is for “Dietrich,” the name of the inventor); it’s a giant vacuum powered by a lawn mower engine that we strap on our backs in the field (see cover photo). Imagine how many times we were heckled for our ghostbuster-like apparatus! We measured “non-crop plant growth” by taking photos of the soil surface along a grid. We’ve combined these various sample techniques and calculated an index of predator diversity and evenness at each farm, which has enabled us to test our hypotheses about each of the environmental factors that might predict predator community structure.
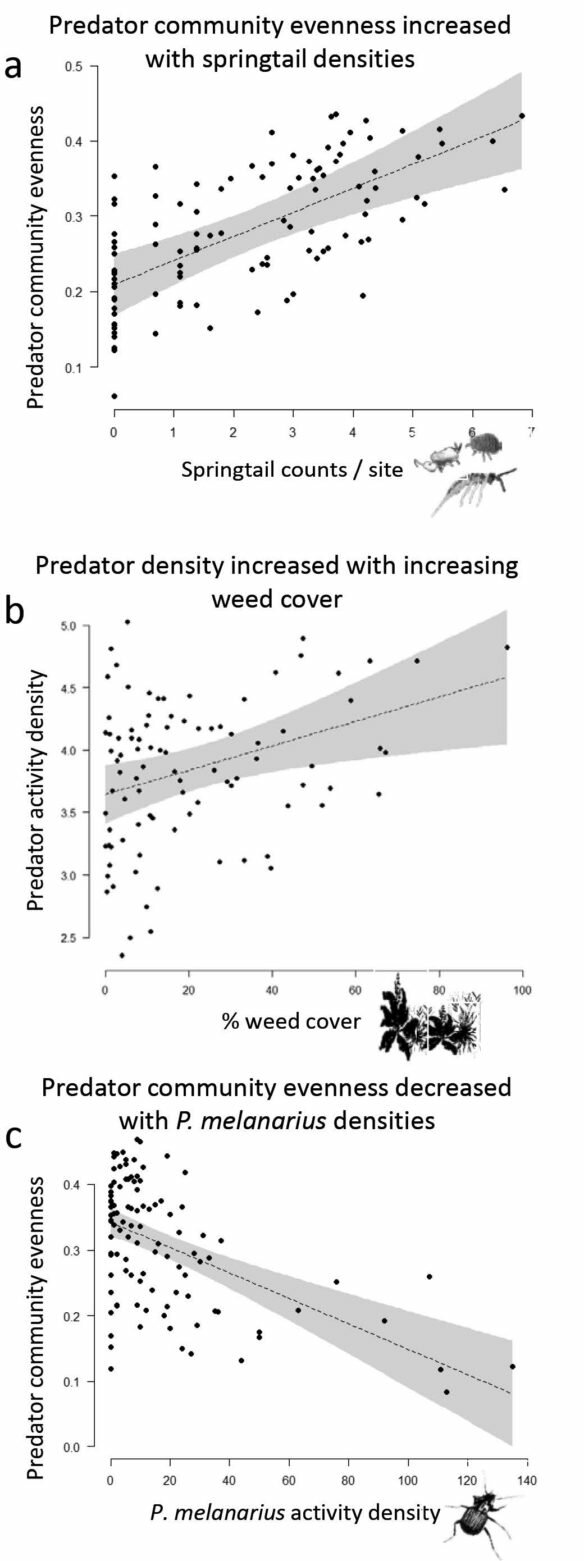 |
| Figure 2. Effects of alternative prey (a), weed cover (b), and a dominant predator (c) on predatory insect communities sampled at more than 50 organic farms. |
Our initial analyses focused on the predator community at the soil surface — they include ground beetles, rove beetles, ants, and spiders. We keyed in on ground predators because they are most likely to prey on flea beetle larvae. Flea beetles are arguably the most hated and difficult vegetable pest to manage in the Pacific Northwest, according to an informal survey I did during a workshop at the last Tilth Conference. Female flea beetles lay their eggs at the base of plants, then the larvae hatch and burrow into the soil, usually feeding on crop roots until they pupate and emerge as adults to wreak havoc on above-ground foliage. It’s during that immature period that they’re most vulnerable to predation, so we’re particularly interested in the role of diversity and community evenness in that flea beetle food web at the soil surface. We found some evidence for our hypothesis that alternative, nonpest prey support stable predator communities in agricultural systems–we counted springtails at each farm, and found that their densities positively predicted predator community evenness (Figure 2a)! Springtails feed on decomposing organic matter in the soil, and are also adversely affected by frequent tillage and insecticide use. So, managing those factors on the farm accordingly may improve the availability of alternative prey, and ultimately support a diverse and even predator community.
Our hypothesis about how non-crop plant availability might support predator community evenness was not clearly supported by our survey results. Weed cover, living mulch and cover crops did not predict community evenness, but they did predict the overall density of predators (Figure 2b). That result was perplexing at first, until we discovered an important underlying detail. It turns out that non-crop habitat is particularly attractive to the most dominant predator in our system, the ground beetle Pterostichus melanarius. P. melanarius in an invader from Europe, and is notorious for being a dominant predator in agroecosystems (an “evenness killer,” if you will). It eats smaller predators, like Bembidion species (B. lampros and B. tetracolum, Figure 1), almost indiscriminately from pests, and can reduce rates of biological control4. Indeed, P. melanarius densities negatively correlated with community evenness across the farms in our survey (Figure 2c), suggesting that this dominant predator compromises predator community structure. Because P. melanarius uses non-crop habitat, effects of cover crops and living mulches on biological control appear more complex than we expected. However, we absolutely recommend incorporating cover crops between rows, as they have myriad other benefits related to soil health, in addition to attracting predators.
Our next steps will be to use more complex statistical models to link interactions between plant, prey, and predator communities to pest suppression. This will help us understand the degree, if any, to which P. melanarius interferes with biological control, and measure the importance of predator community evenness in the field, where it all counts. For now, to conserve stable and even predator communities, we recommend management practices that limit damage to alternative prey populations, like springtails. By reducing tillage and applying insecticides only where/when absolutely necessary, you are more likely to have a healthy resident population of predators available to respond when pests attack.
References
1. Root, R.B. (1973) Organization of a plant-arthropod association in simple and diverse habitats: the fauna of collards (Brassica oleracea). Ecological Monographs 43: 95-124. http://onlinelibrary.wiley.com/doi/10.2307/1942161/full
2. Crowder, D.W., Northfield, T.D., Strand, M.R., and Snyder, W. E. (2010) Organic agriculture promotes evenness and natural pest control. Nature 466: 109-112. http://www.nature.com/nature/journal/v466/n7302/abs/nature09183.html
3. Finke, D.L., & Denno, R.F. (2002) Intraguild predation diminished in complex-structured vegetation: implications for prey suppression. Ecology 83: 643-652. http://onlinelibrary.wiley.com/doi/10.1890/0012-9658(2002)083%5B0643:IPDICS%5D2.0.CO;2/full
4. Prasad, R.P., & Snyder, W.E. (2006) Polyphagy complicates conservation biological control that targets generalist predators. Journal of Applied Ecology 43: 343-352. http://onlinelibrary.wiley.com/doi/10.1111/j.1365-2664.2006.01129.x/full
Carmen Blubaugh, Sabrina Judson and William Snyder, Washington State University Department of Entomology
